
SIGEP
Geological and Paleontological Sites of Brazil
- 033
ROCAS
ATOLL,
SOUTHWESTERN EQUATORIAL ATLANTIC, BRAZIL
Date:17/09/1999
Ruy Kenji Papa de Kikuchi
kikuchi@ufba.br
Departamento de Ciências
Exatas, Universidade Estadual de Feira de Santana
BR-116, Km 3 s/n, Campus Universitário, Feira de Santana, Bahia, Brazil
CEP 44.031-460.
© Kikuchi,R.K.P. 1999.
Rocas Atoll, southwestern equatorial Atlantic, Brazil. In: Schobbenhaus,C.; Campos,D.A.; Queiroz,E.T.; Winge,M.; Berbert-Born,M. (Edit.) Sítios Geológicos e Paleontológicos do Brasil.
Available on line 17/09/1999 at the address http://www.unb.br/ig/sigep/sitio033/sitio033english.htm [Actually
https://sigep.eco.br/sitio033/sitio033english.htm]
[SEE
PRINTED CHAPTER IN PORTUGUESE] (The above bibliographic reference of author copy rights is required for any use of this article in any media, being forbidden the use for any commercial purpose)
|
ABSTRACT
Rocas is the first marine
protected area created in Brazil. It is a Biological Reserve and therefore the
only human activity allowed there is scientific research. It is an ellipsoid
atoll with an internal area of about 7.5 km2. Its largest axis (E-W)
is 3.7 km long, and the shortest (N-S) is 2.5 km long. An algal ridge limits the
reef flat, that is dominated by a coralline algae-vermetid gastropods
association growing as small linear ridges. In the reef front (in some grooves),
in the pools and in the lagoon, corals (Siderastrea
stellata, Montastrea cavernosa and
Porites sp) are found. Seismic
refraction profiles revealed the presence of two subsurface strata. A 11.6 m
long drill core on the western part of the reef, with the recovery rate of 40%,
shows that the Holocene sequence of Atol das Rocas was primarily built by
coralline algae and, subordinately, by corals, along with some encrusting
foraminifer Homotrema rubrum and
vermetid gastropods. The reef growth began before 4.8 ky BP with the accretion
rate varying from 1.5 to 3.2 m/ky. Subaerialy exposed old reef spits, elevated
above tidal range, and a beachrock cliff in one of the cays present in the atoll
are evidences of a equal to or higher than present sea level in Rocas, earlier
in the Holocene. Low degree of competition for space and low grazing pressure
may be the ecological reasons that promoted such a strong growth of coralline
algae in Rocas.
INTRODUCTION
Rocas is a geomorphologic site because it is the only atoll in Southwestern
Atlantic, and one of the smallest in the world. It was discovered in 1503 due to
the sink of Gonçalo Coelho sailboat (Rodrigues
1940)
. Thus, from this very first appearance in the nautical literature, its
low lying profile with only two cays and some rocks outcropping from sea had
always had the meaning of hazardous place.
Rocas
is a geologic site because as a reef it is a carbonate deposit that results from
the building activity response of benthic organisms to environmental parameters
such as light availability, hydrodynamics and relative sea level fluctuation.
Rocas
is a paleontological site because it was built dominantly by coralline algae,
and only secondarily by corals. This is an important fact because it is
generally accepted that coralline algae do not have potential to erect or to be
a primary builder of reefs in the Quaternary (Macintyre
1997)
.
At
last, but not least, Atol das Rocas is an ecological sanctuary because it is
home to a great number of migratory and resident sea birds, which rest, nest and
feed there. Among the most abundant birds in the atoll, there are the brown
tern, the brown and masked boobies and the frigate birds. Furthermore, like
Fernando de Noronha, it is a place of intense nesting activity of green turtles
and feeding activity for the hawksbill turtle. Around Atol das Rocas on the top
of the seamount platform there is a great quantity of commercial fishes.
Mollusks and crustaceans occur in great abundance, as well. Lobsters, as an
example, was one of the causes for the intense predatory fishing activity in the
atoll waters in the past.
The
purpose of this paper is to present an overview description of Atol das Rocas,
based mainly on Kikuchi (1994)
and Kikuchi & Leão (1997)
. Data on its composition and structure will be presented, showing that
the encrusting coralline algae were its primary reef builders. The reasons why
coralline algae predominate in such a conspicuous manner is discussed. I also
present evidences of a former sea level position in the Holocene equal to or
higher than the present sea level. This reinforces the general pattern of the
Brazilian coastal sea level curve that exhibits a transgressive period until
about 5.1 ky BP and a sea regression since that period. A brief comment on the
conservation status of the atoll close this chapter.
ENVIRONMENTAL
SETTING
The atoll is built on the western side of the flat top of a seamount within the
Fernando de Noronha Fracture Zone (Figure 1). It is located about 260 km east of
the city of Natal, in Northeastern Brazil, and 145 km west of the Fernando de
Noronha Archipelago (between 3°45’S and 3°56’S and 33°37’W and 33°56’W).
The lighthouse is located at 3
51’30”S and 33
49’29”W.
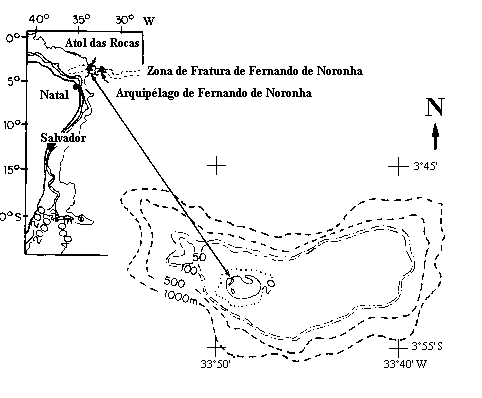
Figure
1: Location map of Atol das Rocas and the biological reserve limits
Kikuchi (1994)
shows, according to data obtained in Instituto Nacional de Pesquisas
Espaciais (INPE), that rainfall is irregularly distributed along the year, with
a monthly average of 860 mm, ranging from 183 mm (April/92) to 2663 mm
(August/92). In this same period, the air temperature ranged from 17.5°C
(April) to 35.8°C (February).
The
data of wind direction indicate that the dominant ESE winds occur along the
whole year, with an average frequency of 45% of the measured days. Between June
and August (southern hemisphere winter) SE winds occur in 35% of the days and
the frequency of E winds is 15% in the same period. Between December and April
(southern hemisphere summer) SE winds and E winds occur in about 20% of the days
with data available. Winds with speed varying between 6 and 10 m/s dominate
along the whole year, but during winter, winds with speed between 11 and 15 m/s
are common. Velocities higher than 20 m/s were registered, more frequently,
during summer.
Tides
are semi-diurnal, with a maximum range of 3.2 m.
The
drift that characterize Atol das Rocas region is the Southern Equatorial
Current, which is originated on the African coast, from the Benguela Current. It
flows to west at a velocity ranging from 30 cm/s to 60 cm/s (Richardson
& McKee 1984; Silveira et al.
1994)
.
According
to Hogben & Lumb (1967)
80% of the waves that were registered in the region where this study
site is included, come from E and 15% from NE. Their periods range from 4 s to 7
s and their heights, from 1 m to 2 m. Melo (1993)
, however, point out that during December and March this behavior can
change, with the occurrence of waves with 15 s to 18 s period and 2 m high waves
coming from the northern hemisphere.
Water
temperature in the outside of the atoll averages 27°C, with minimum value of
25.5°C and maximum value of 28°C. In the inner reef, water in the pools can
reach 39°C. Available data of salinity, indicate that it averages 37 salinity
units (su), varying from 35 su to 42 su. Some data on pH, measured during a
couple of days during the summer of 1991, in the inner part of the reef, show
values varying from 5 (at night) to 11 (at noon) (Kikuchi
1994)
.
Water
visibility during good weather conditions is greater than 20 m. A good
visibility is evidenced in the TM/LANDSAT image (Figure 2), where, in the blue
channel, it is possible to identify bottom features in depths up to 30 m.
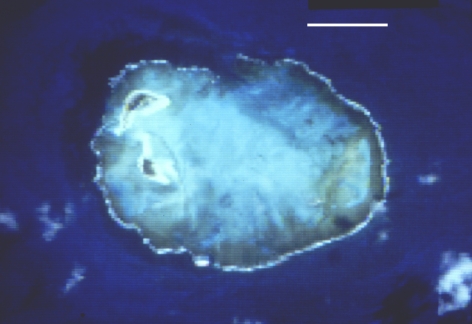
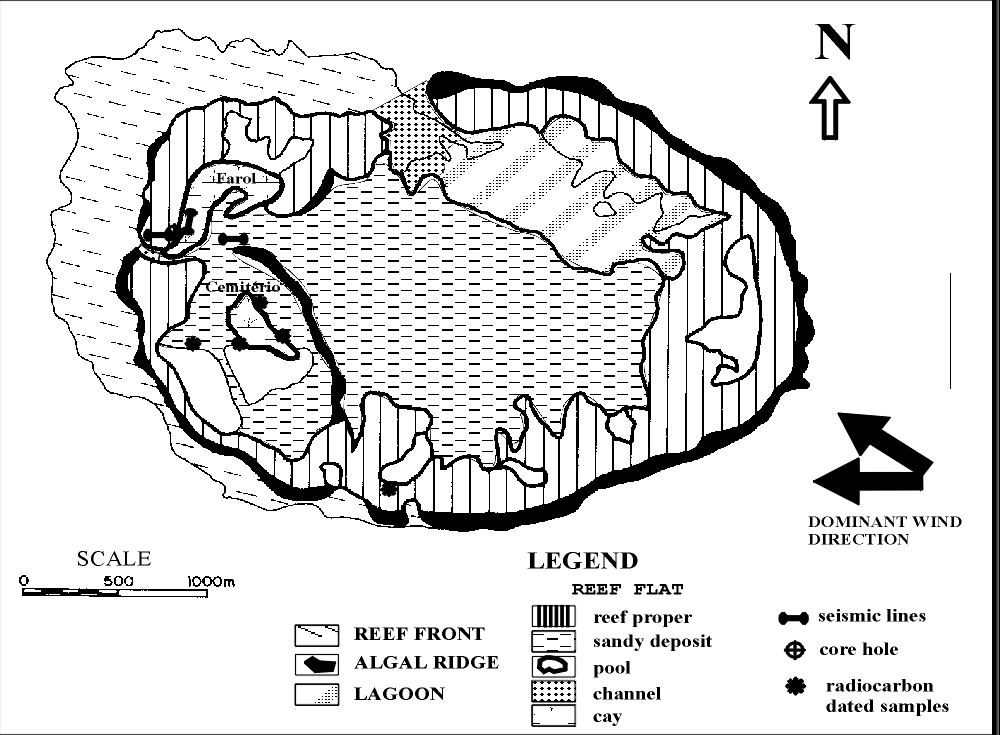
Figure 2: A. TM/LANDSAT satellite image from Rocas atoll, blue band. The scale
bar (white) is 1 km long.
B: Geomorphology map of Atol das Rocas.
HISTORICAL OVERVIEW
The existence of Atol das Rocas was first registered in the XVI Century
Cantino’s Map (Andrade
1959)
. The first detailed map of the atoll apeared in 1852, by Leutnant
Phillip Lee (Rodrigues
1940)
. In that time it was still named Rocas shoal (Baixo das Rocas) or Cabras
Shoal (Baixo das Cabras). It was in the bathymetric chart by Comander Vital de
Farias in 1858 that Rocas was first described as an atoll (Rodrigues
1940)
. Léry (1980)
was the first naturalist to give a brief and faint description the
atoll. It was only in Andrade (1959)
that Rocas received a deeper scientific description. Among other things
he described the morphology of the reef flat, the lagoon, of some pools, the old
reef spits and of the beachrock that occurs in Cemitério islet. These two
latter features were considered indications of a former higher relative sea
level in the atoll and correlated with evidences of higher sea level in the
Holocene found in the Brazilian coast in Pernambuco State (Andrade
1959)
. Thus, Holocene sea level fluctuations were considered of primary
importance on the build-up process of Rocas. Due to the low occurrence of corals
on the reef flat, Andrade (1959)
suggested that Rocas was entirely built by coralline algae (called by
the author as Lithothamnium).
The
controversy about the reef's composition and its classification as an atoll
began with the paper by Vallaux (1940)
. He points out that the reef is composed by coralline algae and that the
lagoon would not deserve this name because of its shallow depths. The issue was
founded on the dispute that occurred in that time between the theories about the
driving mechanism of reef evolution. Darwin's ideas about the 3 successive
stages of reef evolution in the Pacific, from fringing reefs to barrier reefs
and finally to atolls, all driven by isostasy was jeopardized by Daly who
proposed an alternative hypothesis. According to this author, eustatic sea level
changes were responsible for carbonate accretion and solution. Thus, high
frequency sea level rise and fall along geologic history were considered the
cause of reef evolution. The first hypothesis implied that carbonate accretion
on atolls would be thick, with deposits at least as old as the early Tertiary,
and the lagoons great depths were taken as evidences of this processes.
According to the second, the glacial-control theory, reefs in general (and
atolls in particular) would be a thin Pleistocene strata since reef substrate
would not have changed its position throughout reef development. Later in this
century, with the results of borings in the Pacific atolls it was proved that
Darwin's ideas were correct as a general model of reef evolution but, at the
same time, it was seen that the Quaternary thickness of reefs was generally thin
and that eustatic sea level change had an important role in reef development.
Thus, the residual blocks of reefs found on the eastern side of the atoll
surface and the beachrocks present on one of the atoll cays, that indicate a
higher sea-level in the Holocene, are not evidences that falsify the
classification of Rocas as an atoll, as pointed out by Andrade (1959)
in support to Vallaux's ideas. Based on its geomorphological aspects, it
will be shown that Rocas is a true atoll, despite its shallow (6 m deep) but,
nevertheless, navigable lagoon.
The
tectonic setting and substrate character of the atoll were considered by Almeida
(1955), who states that Rocas atoll and Fernando de Noronha Arquipelago belonged
to an alignment of seamounts, which is a branch of the meso-oceanic chain. Bryan
et al. (1973) shows evidences of the
continuity of this alignment through the continental margin in the Rio Grande do
Norte State and into the Brazilian coast and hinterland in the Ceará State.
Gorini (1981) confirms the morphology of this sea-mountains alignment and named
it as Fernando de Noronha Fracture Zone. According to the last author, this
fracture zone is a counterpart of the Jean Charcot Fracture Zone, in the eastern
margin of the South Atlantic Ocean. Cordani (1970) obtained ages in Fernando de
Noronha rocks spanning from 12 to 1,8 m. y. B.P. He adds, though, that the
volcanic activity of the Archipelago might have begun earlier, around 39 m. y.
B.P. Thus, sitting to the west of Fernando de Noronha, Rocas' sea-mount must
have begun to form earlier. Nevertheless the volcanic activity in the seamount
do not necessarily stopped much earlier.
Andrade
(1960) shows that the beachrock grain size is similar to that of the islets'
sediment. Coutinho & Morais (1970) study samples collected on the bottom
surface of Rocas and Fernando de Noronha and observe that it is “biogenic”
carbonate sand, composed mainly by coralline algae (Melobesioidae, Family
Coralinaceae), plates of the green calcareous algae Halimeda
and benthic foraminifers (mainly Amphistegina
radiata e Archaias sp). Tinoco (1972)
studies the foraminifers fauna of bottom surface sediments of Rocas and
found that Amphistegina radiata and Peneroplis
proteus dominates the sediment to
depths greater than 45 m but Archaias
angulatus is more abundant near the atoll and inside it.
SITE DESCRIPTION
Geomorphology
Atol das Rocas is a reef that developed on the flat top of a seamount (Figure
1), where depth ranges from 15 m to 30 m. It is an atoll with elliptical shape,
opened on its western and northern parts. Its greater axis, oriented E-W, is
about 3.7 km long and the minor axis, oriented N-S, is about 2.5 km long (Figure
2). Despite its small dimensions, the reef front, reef flat and a lagoon can be
clearly distinguished and subdivided into discrete features, such as reef front,
reef flat and lagoon (Figure 2A).
The
reef front has two distinct features: on the windward side (eastern and
southeastern side), it is an abrupt, nearly vertical wall, from the reef edge to
depths of about 10 m where a talus deposit occurs on its foot down to depths of
15 m. At this level, there is a flat surface colonized by fleshy and coralline
alga, corals and sponges, which extends up to 1 km to the east and south of the
atoll. In spite of the fact that this surface is colonized dominantly by green
and brown algae, with little or no sediment accumulation, colonies of Mussismilia
hispida and Millepora alcicornis,
together with sponges and rhodoliths are found on it. This is possibly the
surface of the substrate of Atol das Rocas. On the lee side a spur-and-groove
system develops from the reef edge to depths of 18 m.
The
reef flat is subdivided in two components: the reef proper and a sandy deposit
(Figure 2B). The reef proper (reef ring) is 100 to 800 m wide, interrupted
occasionally by pools and opened by two channels in the leeward side of the
atoll, one on the western side and another at the northern side. These two
channels divide the reef proper into the windward arch and the leeward arch.
In
the reef flat there are features like channels, pools and sand cays. The reef
proper or the reef ring is the outer reef flat and contour the sandy deposit and
the lagoon. The surface of the reef proper is composed of linear ridges of
encrusting coralline algae associated with vermetid gastropods, most of which
are approximately parallel to the reef edge (Figure 2B). Branching coralline
algae (Jania sp., Amphiroa sp.),
green and brown macroalgae also grow on these linear ridges. Residues of a
former higher reef building (Figure 3) appear at the windward arch of the atoll,
more frequently on the eastern part of the reef flat, but can be found also on
its southwestern part. This feature, named old reef spits or féo
in the literature (Battistini et al. 1975)
stands up 2 to 3 m above the surface of the reef flat, in Rocas (Figure
3). Notches on the spits feet (Figure 3) indicate that the mean high water
springs (MHWS) is about 0.5 m above the reef flat surface. They are composed
mainly of encrusting coralline algae. Vermetid gastropods and the foraminifer Homotrema
rubrum occur as subordinate components. The sandy deposit corresponds to a
greater part of what Andrade (1959)
calls as “very shallow lagoon”. It is composed mainly by a coralline
algae debris, medium to fine sand-grained. More than 50% of the fragments are
algal debris, with foraminifers tests and mollusk fragments appearing
subordinated (the mean frequency reaches about 10% each). In some places there
appear discontinuous and asymmetric ripples, produced by tidal flow on the reef
flat.

Figure
3: Photography of reef flat in the windward arch, E part of the atoll
On the edge of the windward arch there is a continuous algal ridge, about 20-30
m wide and nearly 0.5 m high. It is exposed above sea level during the low
tides. This is higher energy environment of the reef due to the fact that the
energy of the dominant wave train is dissipated on the reef edge. The algal
ridge is absent from the edge of the leeward arch, in the northwest side or
Farol cay.
A
shallow lagoon, with maximum depth of 6 m, is seen on the northeastern side of
the reef. It communicates with open sea through the northern channel (Figure 2),
called Barreta Grande. This channel is about 100 m wide and almost 100 m long.
Its depth varies from 6 m on its inner part to 10 m on its outer side. This
channel resembles a set of meandering small channels due to the growth of
several pinnacles in it (Figure 4). The corals Montastrea
cavernosa, Siderastrea stellata
and Porites sp grow on the walls of
the pinnacles. The bottom surface of the spaces between the pinnacles is
composed of gravelly sand sediment. The flow through this channel is tidally
controlled. The western channel on the other hand, is much smaller and short
(less than 50 m in each dimension). The flow through this channel is constantly
to outside of the atoll.

Figure
4: Passage between two pinnacles in Barreta Grande. Depth is about 6 m and
bottom surface is made of gravel
Several
pools occur on the reef proper (Figure 2B). They are 3 m deep at maximum, during
the low tides and can attain up to 400 m in length. This is the case of the
Turtle pool, on the eastern side of the atoll. The borders of the pools develop
overhangs and the coalescence of several individual pinnacles can also be seen
(Figure 5). This is an evidence of the process that originated the reef ring. In
the greater pools, isolated pinnacles can be seen as well. The bottom of the
pools are covered with sand.

Figure 5:
Pool on the south part of the atoll. Note the flat top of the pinnacles at water
surface level on the central part of the photo and the transition to the reef
flat.
There are
two sand cays on the western part of the atoll (Figure 2). The southern cay is
called Cemitério and has a cross bedded beachrock cliff 1.5 m high, on its
northeastern side. The height of this cay is about 2 m above the reef flat
level. A lighthouse was built on the northern cay called Farol. This cay is
about 3 m higher than the reef flat level and there is not beachrock developed
on it.
Rocas is
composed of zones frequently found in the Caribbean atolls despite its reduced
dimensions and formation of an semi-closed ring (Figure 2B, Table 1). This
latter characteristic is not a common feature found in the described atolls
(Stoddart 1965). However, comparing Rocas with other Atlantic atolls, many
similarities in the general morphology of the reef can be seen, as shown in
Table 1. Examples of this latter are the slope of the fore reef, that can be
compared to that found in Alacrán reef (Kornicker and Boyd 1962); the width of
the reef proper (ring), that is of the same order found in the Nicaraguan atolls
(Milliman 1969); and the thickness of the Holocene section and the physiographic
origin, that can be matched with that of Hogsty reef (Milliman 1967).
Table
1: Comparison of morphologic characteristics
between Rocas and the other Atlantic atolls
|
|
ROCAS
(Kikuchi
1994)
|
HOGSTY
(Milliman 1967)
|
ALACRÁN
(Kornicker &
Boyd 1962)
|
NICARAGUA
(Milliman 1969)
|
BELIZE
(Stoddart 1962)
|
|
Diameter (km)
|
2.5 x 3.7
|
5 x 9
|
11 x 22
|
3.5x8.5 to 16x32
|
7.5x35 to 16x49
|
|
Area (km2)
|
7.5
|
±40
|
259
|
25 to 260
|
203 to 530
|
|
Seamount top slope
|
0.2° to 0.15°
|
31° to 61°
|
0.02°
|
?
|
14° to 18°
|
|
Depth of lagoon (m)
|
0 to 6
|
6 to 8
|
15
|
10 to 20
|
6 to 43
|
|
Reef ring width
(km)
|
0.2 to 1
|
2
|
2.5
|
0.5
|
|
|
Holocene thickness
(m)
|
> 11.4
|
18
|
33.5
|
?
|
?
|
|
Continuity
|
closed
|
open leeward
|
open leeward
|
open leeward
|
open leeward
|
|
Physiografic
province
|
seamount
|
seamount
|
continental shelf
|
banks
|
platform banks
|
Reef structure and
composition
The results of the seismic survey, allowed the detection of 3 strata. Kikuchi (1997)
published a reevaluation of the data presented in Kikuchi (1994), in
which the mean velocities of each strata were the following:
v0
= 0.33m/ms
v1 = 2.50m/ms
v2 = 4.70m/ms
from the
shallower to the deeper strata.
Thus, the
layers thickness are:
z0
= 1.7m
z1 = 10.0m
were z0
and z1 summed up together are thickness of the Holocene section of
the reef. The bedrock (v2) upper limit depth in the investigated
site, thus, has a minimum value of 11.7m (Figure 6).
The upper two seismic layers of the reef sequence can be
identified as only one layer of Holocene reefrock, based on data from the core
(Figure 6). The low velocity that defined the upper seismic layer, could
represent the occurrence of a cap of reef with water and air in its pores,
resulting from the low level of the tide at the moment of the seismic registers.
The Holocene section of Rocas atoll was almost completely cored. The ages
yielded by coral skeletons confirmed the Holocene age for this section. The core
hole drilled to a depth of 11.69m, with a recovery rate of 40%. Encrusting
coralline algae was the most abundant organism found in the core samples,
forming more than 60% of the recovered rock. This prominent role of coralline
algae in the building up of reefs is a common feature of Brazilian reefs, such
as seen in Abrolhos and in the northern coast of Bahia. A similar characteristic
is observed in reefs of Bermudas (boilers), studied by Ginsburg (1973)
, and also in the reefs of Antilles, studied by Adey & Burke (1977)
. Coral skeletons appeared only subordinately. They were fragments or
small colonies of the species Siderastrea stellata, Favia
gravida, Mussismilia hispida, Agaricia
sp and Porites sp and made up about 10% of the rock. Vermetid gastropods
and the encrusting foraminifer Homotrema
rubrum are responsible, each of them, for about 6% of the core and occur
associated with the coralline algae.
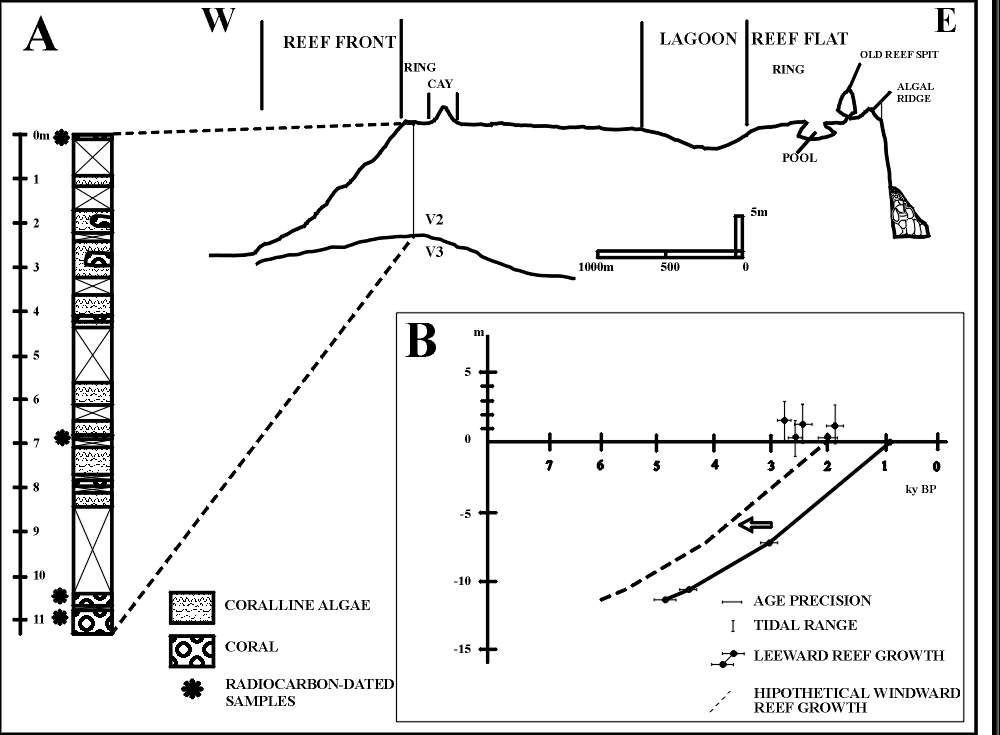
Figure
6: Schematic W-E profile of Rocas atoll, with interpretation of seismic survey.
The core hole is represented on the profile, together with its schematic
composition. The ages of the coral skeletons from the core are also plotted
The
pre-Holocene substrate of Rocas can be compared to the volcanic rocks that
appear in the Fernando de Noronha Archipelago (ultramafic to intermediate
volcanic rocks, according to Almeida, 1955
). The seismic velocity that characterize this lower layer is of the same
order of those from the basalts described in the subsurface of Bikini atoll
(Dobrin et al. 1949; Raitt 1954), in
Kwajalein and Sylvania guyot (Raitt 1953) e no atol Eniwetak (Raitt 1957). This
basal section (Figure 6) may be Tertiary, considering the ages obtained in the
volcanic rocks of Fernando de Noronha Archipelago dated in Cordani (1970).
Reef accretion and sea
level position
The 14C ages obtained from the core are 4.86 ky BP at the depth of
11.2 m, 4.41 ky BP at the depth of 10.5 m, 3.06 ky BP at the depth of 7 m and
0.84 ky BP at the surface (Figure 6, Table 2), and they indicate that the 2.5
m/ms layer corresponds to a Holocene reef sequence. Consequently reef growth, at
the cored site, must have begun at about 5 ky BP and grew up to present sea
level with an average accretion rate of 2.8 m/ky (from 1.5 m/ky to 3.2 m/ky,
Table 3). The age of a skeleton of S.
stellata, of 2.02 ky BP, found in life position on one small old reef spit
on the southwestern part of the windward arch, 0.5 m above the level of the reef
flat surface, indicates that parts of the reef reached the present level of the
sea at about 2.0 ky BP. The dates yielded by the beachrock ranging from 1.91 ky
BP to 2.83 ky BP (Table 1), are concurrent with the age of the coral on the reef
spit, what reinforces the hypothesis that some parts of the reef have already
reached the present position of sea level between 3.0 ky BP and 2.0 ky BP.
Table
2: 14C ages of coral skeletons (Ss = Siderastrea
stellata, Fg = Favia gravida) and
mollusk shells (mol) from the reef surface, the core samples and the Cemitério
island beachrock.
|
Location
|
Material
|
Lab no.
|
Conventional 14C
age
(ky BP)
|
|
reef front, 10 m
deep, surface
|
Ss
|
Bah-1758
|
recent
|
|
reef flat, 0.5 m
above surface
|
Ss
|
Bah-1759
|
2.02 ± 0.16
|
|
reef flat surface
|
Ss
|
Bah-1803
|
0.94 ± 0.14
|
|
core top
|
Ss
|
Bah-1801
|
0.84 ± 0.14
|
|
core,
7.0 m deep, leeward ring
|
Ss
|
Bah-1806
|
3.06 ± 0.18
|
|
core, 10.5 m
deep, leeward ring
|
Ss
|
Bah-1807
|
4.41 ± 0.20
|
|
core, 11.2 m
deep, leeward ring
|
Ss
|
Bah-1808
|
4.86 ± 0.21
|
|
beachrock, 0.5 m
above reef flat
|
Fg
|
Bah-1800
|
2.63 ± 0.15
|
|
beachrock, 1.5 m
above reef flat
|
Ss
|
Bah-1796
|
1.91 ± 0.15
|
|
beachrock, 1.8 m
above reef flat
|
mol
|
Bah-1798
|
2.51 ± 0.17
|
|
beachrock, 2.0 m
above reef flat
|
Fg
|
Bah-1797
|
2.83 ± 0.16
|
The oldest 14C date obtained from the core (4.8 ky BP, Table 2) may
not represent the very beginning of reef development in Rocas. There may be a
difference in the reef age between the windward arch and the leeward arch, where
the core was bored. This difference is suggested by the following: i) the
continuos and well developed algal ridge on the windward arch, contrary to what
happens in the leeward arch, where the algal ridge is a feeble and discontinuous
feature; ii) the presence of reef residues (old reef spits) higher than
today’s sea level, on the windward arch (Figure 3); iii) the ages of the
skeletons collected on the reef flat, on the old reef spit and in the beachrock
indicate a possible difference of about 2.0 ky (Table 1) between the time when
the reef surfaced at the windward part and the time when it surfaced at the
leeward part. Considering that the reef accreted at the same average rate (2.8
m/ky, Table 3) in the two arches, reef growth should, thus,
have begun earlier on the windward arch, say at about 6.0 ky BP (Figure
6). It developed as an open atoll and the heights of the old reef spits indicate
that at 2.0 ky BP the reef should have reached up to 3 m above the today reef
flat surface (Table 1). The leeward arch began to develop at about 5.0 ky BP,
with an accretion rate increasing from 1.5 m/ky to 3.2 m/ky. Consequently, the
reef may have attained its semi-closed shape only recently, after the leeward
arch reached its present level, after 1.0 ky BP (Table 2).
Table
3: Reef growth rates, calculated from the ages of the coral skeletons obtained
in the core. Depth calculated with reference to reef flat level.
|
Interval
(m)
|
Rate
(mm/year)
|
|
0
- 7,0
|
3,2
|
|
7,0
- 10,5
|
2,4
|
|
10,5
- 11,2
|
1,5
|
|
average
|
2,8
|
Reef building flora and
fauna
The reef surface is mainly covered by soft algae and an association of living
coralline alga and vermetid gastropods. A study of the corallines by Gherardi (1999)
indicates the occurrence of the genera Porolithon,
Lithophyllum and Sporolithon,
and Lithoporella among encrusting
coralline algae. Massive corals, such as Siderastrea
stellata, Montastrea cavernosa and
Porites sp, only occur in protected
areas, mainly in the lagoon, within the pools and in some grooves of the reef
front. (Echeverría
et al. 1997)
published a list of cnidarians of Rocas in which, among the reef
building corals, Madracis decactis, Agaricia
agaricites, Porites astreoides, Porites branneri, Favia
gravida, and Mussismilia hispida
are cited, besides those species mentioned before. The almost absolute dominance
of Siderastrea stellata among the
hermatypic coral species is a striking characteristic of Rocas.
Two
aspects about the diversity of reef building organisms must be assessed. First
of all, it is the present dominance of coralline algae, which occurred along
whole development of the reef. Second, among reef building corals there is a
conspicuous dominance of a unique species, Siderastrea
stellata.
Although
it is presently recognized that coralline algae has a limited role in reef
building (Macintyre 1997)
due to ecological and environmental constraints and to their low
accretion rate (see Steneck 1986 and references wherin), a 11 m thick of reef
rock formed primarily by corallines accreted in a quite rapid average rate at
Rocas (2.8 m/ky). Figueiredo (1997)
shows that in Abrolhos also coralline algae growth is more elevated than
in reef environments elsewhere. Besides high hydrodynamic energy, low
inter-specific competition and a low degree of herbivory are conditions that
favor coralline algae development (Steneck
1997)
. It is possible that all these three conditions are met in Rocas. The
few number of coral species and the low cover rate of the reef surface by corals
that characterize Rocas, may result in a lower degree of competition for space
between corallines and corals than that found in other reef sites, and this may
have resulted in the enhancement of the coralline algae development. On the
other hand, although the biomass of grazers at Rocas seems to be equivalent to
the biomass of grazers in other reef sites in Brazil or in the Caribbean region
only one genera of coralline algae grazer parrot fish (Sparisoma)
is found at Rocas (Rosa
& Moura 1997)
. The fishes of the genera Sparisoma have weaker mouth muscles than those of the Scarus
genera, the commoner reef grazer, which are a more powerful coralline algae
grazer than Sparisoma. This difference in the composition of the fish fauna may
have contributed, also, to the enhancement of the coralline algae growth and
preservation since the grazing activity is one of the most important ecological
constraint to the development of coralline algae (Steneck 1986)
.
Considering
the dominance of Siderastrea stellata, Echeverría (1997)
suggest that the tolerance to environmental conditions such as high wave
energy and to high temperature variations may be the reason for the success of
this species in the atoll. The contribution of Geomorphology must be stressed,
as well. Due to the reduced dimensions of Rocas, water warmed during the low
tides (39°C to 40ºC), may influence the lagoon as well. The lagoon, besides
the pools is where corals can grow in profusion in the atoll.
CONSERVATION STATUS
Atol das Rocas is a biological reserve that belongs to the State of Rio Grande
do Norte. It is the first marine protected area established in Brazil, enforced
by Federal Act n° 83.549 of July 5th, 1978. In the National System
of Conservation Units (Sistema Nacional de Unidades de Conservação – SNUC),
biological reserve is a protected area category created for full conservation of
biodiversity. No recreational activity nor resources exploitation is admited
within these areas. However, visiting for scientific research and educational
purposes is allowed in special cases, with prior allowance of Brazilian
Institute of the Environment and of the Natural Renewable Resources (Instituto
Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis – IBAMA ).
The
Biological Reserve of Atol das Rocas includes not only the reef itself but all
seamount top limited by the 1000 m isobath. Its total area sums up to 360 km2.
Though it was created in 1978, it was not until 1990 that the conservation
activities began for real. The first reserve station camping was established
under the auspices of Marine Turtle Foundation (Fundação Pró-TAMAR) and of
Manatee Project (Projeto Peixe-Boi Marinho – IBAMA).
It
takes generally 26 hours to arrive the atoll, from the city of Natal (Rio Grande
do Norte). Research teams composed generally of 8 people shift every 25 days.
Each team is composed of 2 park rangers (from IBAMA) and 6 more people
(scientists, students and volunteers). Communication with the continent are made
with the use of SSB and VHF radio systems.
ACKNOWLEDGMENTS
I
would like to thank the organizers of this book for the invitation and the
opportunity to write about a beloved object. Also, I’d like to thank Prof. Dr.
Zelinda M. A. N. Leão for opening the doors for me to the study of the
carbonate environment. Mr. Gilberto Sales, former chief, and Ms. Zélia Brito,
present Chief of Reserva Biológica do Atol das Rocas gave me the opportunity of
developing the fieldwork and helped during the field work in Rocas. The
TM/LANDSAT image was acquired in Instituto Nacional de Pesquisas Espaciais
(INPE).
BIBLIOGRAPHIC REFERENCES
Adey, W.; Burke, R.B.
1977. Holocene bioherms of Lesser Antilles - Geologic control of development.
In: S.H. Frost; WEISS, M.P.; SAUNDERS, J.B. eds.
Reefs and related carbonates -
Ecology and Sedimentology.Tulsa, American Association of Petroleum
Geologists. p. 67-81.
Almeida, F.F.M. 1955. Geologia
e Petrologia do Arquipélago de Fernando de Noronha. Rio de Janeiro,
DNPM/DGM. 181 p.
Andrade, G.O. 1959. O
recife anular das Rocas (Um registro das recentes variações eustáticas no Atlântico
equatorial). Anais da Associação dos Geógrafos
Brasileiros, XI: 29-61.
Battistini, R.,
et al. 1975. Éléments de terminologie récifale Indopacifique. Téthys,
7: 1-111.
Bryan, G.M.; Kumar, N.;
Castro, P.J.M. 1973. The north Brazilian ridge and the extension of equatorial
fracture zones into the continent. In:
CONGRESSO BRASILEIRO DE GEOLOGIA, XXVI, Belém, Anais...,
SBG-Norte, p. 133-144.
Cordani, U.G. 1970. Idade
do vulcanismo do Atlântico Sul. Boletim
do Instituto de Geciências e Astronomia - USP, 1: 9-76.
Coutinho, P.N.; Morais,
J.O. 1970. Distribución de los sedimentos en la plataforma continental
norte-nordeste del Brasil. In: SYMPOSIUM ON INVENTORY OF RESOURCES OF THE
CARIBBEAN SEA AND ADJOINING REGION. Curaçao, 1970. Proceedings...,
p. 273-284.
Dobrin, M.B.; Perkins Jr,
B.; Snavely, B.L. 1949. Subsurface constitution of Bikini Atoll as indicated by
a seismic refraction survey. Bulletin of
the Geological Society of America, 60:807-828.
Echeverría, C.A.; Pires,
D.O.; Medeiros, M.S.; Castro, C.B. 1997. Cnidarians of the Atol das Rocas. In: INT. CORAL REEF SYM, 8th, Panama, 1996. Proceedings... ISRS. V. 1, p. 443-446.
Figueiredo, M.A.O. 1997.
Colonization and growth of crustose coralline algae in Abrolhos, Brazil. In: INT. CORAL REEF SYM, 8th, Panama, 1996. Proceedings... V. 1, p. 689-694.
Gherardi, D.F.M.; Bosence,
D.W.J. 1999. Modeling of the ecological succession of encrusting organisms in
recent coralline-algal frameworks from Atol das Rocas, Brazil. Palaios,
14: 145-158.
Ginsburg, R.N.; Schroeder,
J.H. 1973. Growth and submarine fossilization of algal cup reefs, Bermuda. Sedimentology, 20: 575-614.
Gorini, M.A. 1981. The
tectonic fabric of the Equatorial Atlantic and adjoining continental margins:
Gulf of Guinea to Northeastern Brazil. In: Asmus, H.E. ed.
Estruturas e tectonismo da margem
continental brasileira e suas implicações nos processos sedimentares e na
avaliação do potencial de recursos minerais.Rio de Janeiro, PETROBRÁS,
CENPES, DINTEP. p. 11-116.
Hogben, N.; Lumb, F.E.
1967. Ocean wave statistics. London, National Physical Lab., Ministry of
Technology. 263 p.
Kikuchi, R.K.P. 1994.
Geomorfologia, Estratigrafia e Sedimentologia do Atol das Rocas
(Rebio-IBAMA/RN). Salvador, 144 p. (Dissertação de Mestrado, Pós-Graduação
em Geologia, Universidade Federal da Bahia).
Kikuchi, R.K.P.; Leão,
Z.M.A.N. 1997. Rocas (Southwestern Equatorial Atlantic, Brazil): an atoll built
primarily by coralline algae. In: INT.
CORAL REEF SYM, 8th, Panama, 1996. Proceedings...
V. 1, p. 731-736.
Kornicker, L.; Boyd, D.W.
1962. Shallow-water geology and environments of Alacran Reef Complex, Campeche
Bank, Mexico. Bulletin of the American
Association of Petroleum Geologists, 46: 640-673.
Léry, J.d. 1980. Viagem
à terra do Brasil. São Paulo, Editora Itatiaia/Ed. Universidade de São
Paulo. 303 p.
Macintyre, I.G. 1997.
Reevaluating the role of crustose coralline algae in the construction of coral
reefs. In: Proc 8th Int Coral Reef Sym. Panamá, 1996, ISRS. V. 1, p.
725-730.
Melo, F., Eloi; Alves,
J.H.G.M. 1993. Nota sobre a chegada de ondulações longínquas à costa
brasileira. In: Simpósio Brasileiro de Recursos Hídricos, X, Gramado, 1993. Anais...,
ABRH, p. 362-369.
Milliman, J.D. 1967. The
Geomorphology and history of Hogsty Reef, a Bahamian atoll. Bulletin
of Marine Science, 17: 519-543.
Milliman, J.D. 1969. Four
southwestern Caribbean atolls: Courtown Cays, Albuquerque Cays, Roncador Bank
and Serrana Bank. Atoll
Research Bulletin, 129:1-26.
Raitt, R.W. 1954.
Seismic-refraction studies of Bikini and Kwajalein atolls. Bikini and nearby
atolls. Part 3. Geophysics. US Geological
Survey Professional Paper, 260S: 507-527.
Raitt, R.W. 1957.
Seismic-refraction of Eniwetak atoll. Bikini and nearby atolls. Part 3.
Geophysics. US Geological Survey Professional Paper, 260S: 685-698.
Richardson, P.L.; McKee,
T.K. 1984. Average Seasonal-Variation of the Atlantic Equatorial Currents From
Historical Ship Drifts. Journal of
Physical Oceanography, 14: 1226-1238.
Rodrigues, O.A.A. 1940. O
Atol da Rocas. Revista Marítma Brasileira,
LIX: 1181-1228.
Rosa, R.S.; Moura, R.L.
1997. Visual assessment of reef fish community structure in Atol das Rocas
Biological Reserve, off northeastern Brazil. In:
INT. CORAL REEF SYM, 8th, Panama, 1996. Proceedings...
V. 1, p. 983-986.
Silveira, I.C.A.; Miranda,
L.B.; Brown, W.S. 1994. On the origins of the North Brazil Current. Journal
of Geophysical Research, 99: 22501-22512.
Steneck, R.S. 1986. The
ecology of coralline algal crusts: Convergent Patterns and Adaptive Strategies. Annual Review of Ecology and Systematics, 17: 273-303.
Steneck, R.S. 1997.
Crustose corallines, other algal fuctional groups, herbivores and sediments:
complex interactions along reef productivity gradients. In:
INT. CORAL REEF SYM, 8th, Panama, 1996. Proceedings...
V. 1, p. 695-700.
Stoddart, D.R. 1965. The
shape of atolls. Marine Geology, 3:
369-383.
Tinoco, I.M. 1972. Foraminíferos
dos bancos da costa nordestina, Atol das Rocas e Arquipélago de Fernando de
Noronha. Trabalhos do Instituto Oceanográfico
da Universidade Federal de Pernambuco, 13: 49-60.
Vallaux, C. 1940. La
formation atollienne de Rocas (Brésil). Bulletin
de L'Institut Océanographique, 37: 1-8.